|
|
||||
|
|
|
|
|
|
|
|
||||
|
|
||||
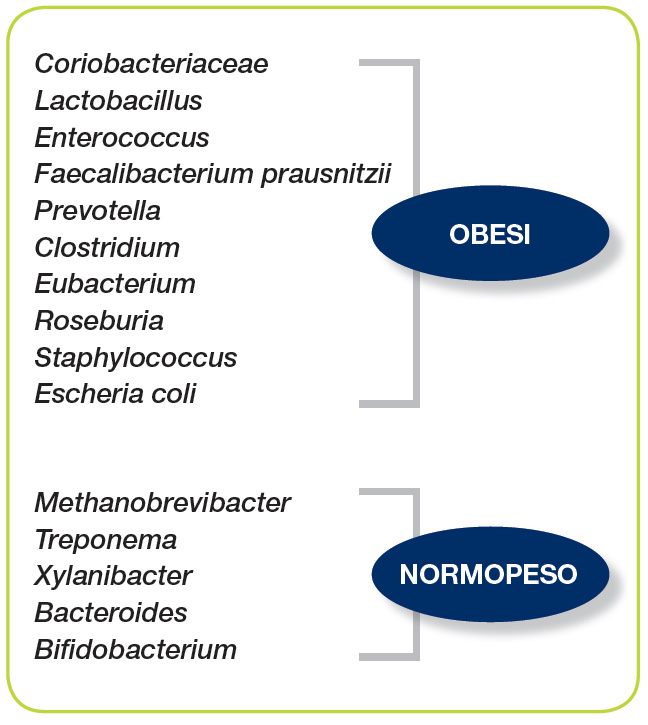
Microbiota intestinale ed obesità L’obesità è una condizione clinica caratterizzata da un eccesso ponderale secondario ad accumulo di tessuto adiposo tale da comportare un indice di massa corporea ≥30 Kg/m2. Diffusa soprattutto nei Paesi industrializzati fino agli anni ’80, l’obesità rappresenta ai giorni nostri una vera e propria emergenza “epidemiologica” mondiale iniziando a diffondersi persino nei Paesi in via di sviluppo, dove, storicamente, erano prioritari i problemi di malnutrizione. È stato calcolato che negli ultimi 30 anni, la prevalenza di obesità negli adulti è aumentata del 75% e che, attualmente, più della metà della popolazione statunitense è in sovrappeso. In media, 1 adulto su 3 è clinicamente obeso e progressivamente il problema sta iniziando a coinvolgere anche le fasce pediatriche della popolazione, guadagnando sicuramente un ruolo prioritario per la salute pubblica. L’obesità, infatti, non solo è una condizione di per sé caratterizzata da ridotta qualità di vita, ma soprattutto è uno dei più importanti fattori di rischio per le patologie metaboliche (diabete mellito ed iperlipemie), le malattie cardiovascolari, l’ipertensione polmonare, l’apnea ostruttiva del sonno, la malattia da reflusso gastroesofageo, le patologie muscolo-scheletriche ed il cancro. Inoltre, l’obesità è associata globalmente ad una mortalità più elevata. Da un punto di vista patogenetico, l’obesità è stata a lungo semplicisticamente inquadrata come la risultante di uno sbilanciamento tra introito calorico e dispendio energetico. Il progressivo aumento del consumo di alimenti ad elevato contenuto calorico e la parallela riduzione dell’attività fisica registrata negli ultimi decenni sicuramente hanno contribuito all’aumento della prevalenza dell’obesità. Tuttavia, l’esistenza di complessi sistemi di regolazione dell’omeostasi energetica, implica che il paradigma sia in realtà molto più complesso di quanto finora era stato ritenuto. L’essere umano si è co-evoluto con una comunità di batteri ed altri microrganismi intestinali che complessivamente formano il cosiddetto “microbiota intestinale”. La composizione del microbiota varia non solo tra individui diversi della stessa specie ma anche nello stesso individuo durante l’arco della vita sotto l’influenza di complesse e dinamiche interazioni tra fattori costituzionali, dieta, stile di vita ed uso di farmaci. Il tipo di evoluzione che subisce il microbiota e il suo genoma influenza profondamente la biochimica dell’ospite e può condizionare la suscettibilità alle malattie. La possibile relazione tra microbiota intestinale e obesità è stata inizialmente ipotizzata sulla base di una serie di studi effettuati dal gruppo di ricerca di Jeffrey Gordon della Washington University. Da questi studi è emerso che il profilo del microbiota intestinale differisce profondamente in base al peso corporeo e che i batteri intestinali sono direttamente implicati nella patogenesi dell’obesità (Figura 1).
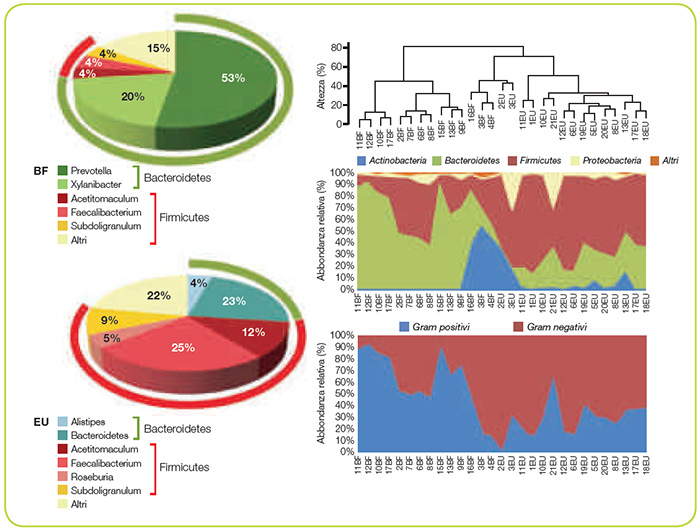
La prima dimostrazione di una specifica alterazione nella flora batterica intestinale tra fenotipo obeso e normopeso è stata ottenuta in topi geneticamente obesi (ob/ob) nei quali, infatti, si osservava una prevalenza di Firmicutes con una parallela riduzione dei Bacteroidetes. Addirittura, nell’animale da esperimento si è dimostrato che il “fenotipo obeso” era trasmissibile con la flora intestinale. Infatti, topi germ-free colonizzati con la flora batterica derivata da animale obeso presentavano un aumento del grasso corporeo di circa il 60% in più rispetto a quelli colonizzati con la flora batterica di un animale normopeso. Analoghi risultati sono stati ottenuti nell’essere umano. Infatti, Ley e collaboratori, nel 2005, esaminando il microbiota intestinale di volontari obesi sottoposti ad un programma di riduzione del peso corporeo, hanno dimostrato che prima dell’inizio della dieta ipocalorica, i pazienti mostravano una prevalenza di Firmicutes, mentre al termine dello studio aumentavano i Bacteroidetes dal 3% iniziale al 15% finale. In accordo con queste osservazioni, in pazienti sovrappeso gravide alla 24ª settimana di gestazione, si è osservata una riduzione del numero dei Bifidobacteri e Bacteroidetes e un parallelo aumento del numero di Firmicutes (es. Staphylococcus) e Proteobacteria (es. Enterobacteriaceae quali Escherichia coli). Inoltre, è stato dimostrato che il cambiamento della flora batterica precede lo stabilirsi del fenotipo obeso e contribuisce al suo sviluppo. La “disbiosi” intestinale comporta un’alterata funzionalità metabolica dell’intestino con differenze individuali significative nel recupero energetico (estrazione calorica dagli alimenti). Infatti, il microbiota intestinale, dotato di enzimi, quali la glicoside idrolasi, non codificati dal genoma umano, contribuisce al recupero di energia da polisaccaridi introdotti nella dieta altrimenti non digeribili. Diverse linee di evidenza suggeriscono che la dieta possa profondamente influenzare la composizione del microbiota intestinale. Un’elegante dimostrazione di quanto la dieta possa essere determinante sull’assetto del microbiota intestinale viene da un famoso studio epidemiologico che ha posto a confronto la composizione della flora batterica intestinale di bambini di un’area europea, a dieta occidentale, con quella di bambini di un’area rurale africana. Questi ultimi presentavano un importante arricchimento in Bacteroidetes e una deplezione dei Firmicutes, con una significativa rappresentanza di batteri del genere Prevotella e Xylanibacter, che sono riconosciuti in grado di codificare geni capaci di idrolizzare la cellulosa e lo xilano. Infatti, i bambini africani avevano anche un più elevato contenuto fecale di acidi grassi a corta catena. Il tipo di flora batterica prevalente nell’area rurale africana in pratica consente di massimizzare l’estrazione di energia dalle fibre introdotte con la dieta, proteggendo tra l’altro dalle infezioni e dall’infiammazione (Figura 2).
Una dieta iperlipidica agisce attraverso la stimolazione della produzione di RELM-β, e della conseguente inibizione dell’espressione di peptidi antimicrobici prodotti a livello intestinale, favorendo in tal modo la predominanza del phylum Firmicutes rispetto al phylum Bacteroidetes. Uno dei ruoli fisiologici espletati dalla RELM-β è quello di stimolare a livello intestinale la produzione di peptidi antimicrobici endogeni (α e β-Defensine). A dimostrazione di ciò, è stato allestito un modello in vitro di cellule CaCo2 (Colonic Epithelial Cells) sottoposte a stimolazione con la proteina ricombinante hRELM-β. Questo studio ha dimostrato che RELM-β riduce l’espressione della ®-Defensina 1, riducendo sensibilmente l’attività antimicrobica del surnatante cellulare. La colonizzazione dei topi germ-free con il fenotipo batterico obeso sopprime l’espressione di un inibitore della Lipoproteina Lipasi Circolante (LPL), il Fasting Induced Adipose Factor (FIAF). L’aumento dei livelli di LPL determina un aumentato uptake cellulare di acidi grassi dai triacilgliceroli associati alle lipoproteine e l’accumulo di trigliceridi negli adipociti. Altri studi dimostrano il ruolo importante esercitato dall’Adenosine Monophoshate activated Protein Kinase. Si tratta di un enzima ereditato dai lieviti in grado di proteggere il topo dall’obesità indotta da un’alimentazione iperglucidica e iperlipidica. Questo enzima si attiva in risposta a condizioni di stress e monitora lo stato energetico cellulare. I topi germ-free rimangono magri nonostante l’elevato introito calorico perché gli alti livelli di AMP-K, determinano un aumento dell’ossidazione degli acidi grassi nei tessuti periferici, una diminuzione dei livelli di glicogeno ed un’aumentata sensibilità all’insulina. Prospettive future Sicuramente l’interazione tra il microbiota intestinale e il proprio ospite diventerà uno dei più importanti temi della ricerca biomedica dei prossimi anni e ci porterà ad una sempre più dettagliata conoscenza del rapporto funzionale tra batteri, metabolismo e obesità. La capacità di modulare la composizione del microbiota intestinale sarà, in un futuro non molto lontano, uno dei cardini per sviluppare strategie di prevenzione e di terapia per le patologie metaboliche correlate all’alimentazione. ----------------------------------------------------------------------------------------------------------------------------------------------------------------
|